- 产地
- 苏州
- 品牌
- 细胞外基质胶
- 型号
- 齐全
- 是否定制
- 是
这种转化细胞接种入正常机体,常能长成块,并侵润正常组织,发生普遍转移。又如上皮细胞分泌胶原蛋白和膜粘蛋白于上皮组织的基底层上,反之,这些蛋白又作为信号“指挥”上皮细胞生长、迁移的方向。在胚胎发育或愈伤再生时,上皮细胞正是沿着基底层发展的。由此可知,调节细胞生长、发育的若干信息正是通过胞外基质传递的。细胞外基质的主要类型及功能:对人类细胞的研究表明,细胞外基质中的纤粘蛋白主要由成纤维细胞、上皮细胞等分泌并附着在细胞表面,其作用是促进细胞对基质的贴附,细胞之间的粘着,细胞内微丝及应力纤维的构建。观察到转化的体外培养的成纤维细胞,表面纤维蛋白量减少,与此相关地细胞形态变圆,与培养基底贴附松弛,胞内应力纤维很大减少,细胞密集,重叠生长。细胞外基质的主要类型及功能:粘连糖蛋白,包括纤连蛋白和层粘连蛋白,有助于细胞粘连到胞外基质上。徐州开封细胞外基质胶

骨膜来源的细胞外基质水凝胶通过早期免疫调节及增强血管和骨生成促进骨修复:骨愈合包括早期炎症免疫调节、血管生成、成骨分化和生物矿化等过程,干预其中的任一过程都可能阻碍骨修复。在复杂和严重的骨损伤部位,大量的促炎因子的存在会诱发促炎反应。长期的促炎反应会阻碍巨噬细胞从M1到M2的转变导致骨再生延迟。因此,在骨损伤早期通过调控M1向M2的转变来适时终止促炎反应是骨愈合成功的前提。近日,浙江大学医学院范顺武和林贤丰教授课题组制备了一种骨膜来源的细胞外基质(PEM)水凝胶,并评价了它们在骨修复过程中不同时期的调节作用南京天津细胞外基质胶大量合成的肾脏细胞外基质取代了肾小球各功能细胞的空间。
细胞外基质的作用:(一)细胞外基质的作用:细胞外基质不只具有连接、支持、保水、抗压及保护等物理学作用,而且对细胞的基本生命活动发挥很全的生物学作用。1.影响细胞的存活、生长与死亡正常真核细胞,除成熟血细胞外,大多须粘附于特定的细胞外基质上才能克制凋亡而存活,称为定着依赖性(anchoragedependence)。例如,上皮细胞及内皮细胞一旦脱离了细胞外基质则会发生程序性死亡。此现象称为凋亡(anoikis,aGreekwordmeaning“homelessness”)。不同的细胞外基质对细胞增殖的影响不同。例如,成纤维细胞在纤粘连蛋白基质上增殖加快,在层粘连蛋白基质上增殖减慢;而上皮细胞对纤粘连蛋白及层粘连蛋白的增殖反应则相反。
细胞外基质的介绍:细胞外基质(extracellularmatrix,ECM),是由动物细胞合成并分泌到胞外、分布在细胞表面或细胞之间的大分子,主要是一些多糖和蛋白,或蛋白聚糖。这些物质构成复杂的网架结构,支持并连接组织结构、调节组织的发生和细胞的生理活动。细胞外基质是动物组织的一部分,不属于任何细胞。它决定结缔组织的特性,对于一些动物组织的细胞具有重要作用。细胞是生物体基本组成单位。绝大多数哺乳类动物细胞之间存在成分复杂的细胞外基质(ECM)。间质基质存在于各种动物细胞之间(即细胞间隙中)。

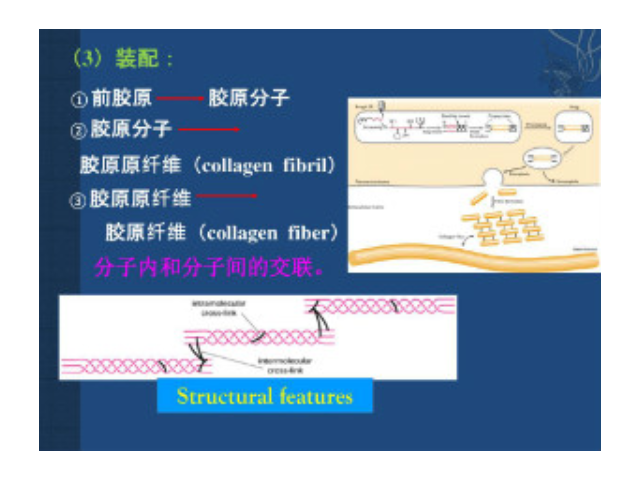
根据胶原的结构和功能可将其分为:纤维性胶原(fibrilformingcollagen)这是较经典的胶原,如Ⅰ、Ⅲ、Ⅴ和Ⅺ型胶原。其肽链长达1000个氨基酸,是结缔组织中含量较丰富的胶原。前胶原三螺旋的端肽被切除后纵向平行排列,其中每个胶原分子纵向稍偏移,相邻的肽链形成共价键交联从而形成微纤维。一般需经前胶原肽酶(procollagenpropeptidase)将羧基端肽去除后才能形成胶原纤维,但是部分胶原可以带有氨基端肽而存在于胶原纤维的表面,以阻止胶原纤维继续增粗,从而继续起到调节胶原纤维直径的作用。系膜细胞病变克制了肾脏纤溶酶的降解活性。昆明细胞外基质胶单价
细胞外基质并非像过去认为的起惰性支持物的作用,或将细胞连接在一起,形成组织、部位。徐州开封细胞外基质胶
细胞外基质层粘连蛋白:(laminin,LN)LN也是一种大型的糖蛋白,与Ⅳ型胶原一起构成基膜,是胚胎发育中出现较早的细胞外基质成分。LN分子由一条重链(α)和二条轻链(β、γ)借二硫键交联而成,外形呈十字形,三条短臂各由三条肽链的N端序列构成。每一短臂包括二个球区及二个短杆区,长臂也由杆区及球区构成。LN分子中至少存在8个与细胞结合的位点。例如,在长臂靠近球区的。链上有IKVAV五肽序列可与神经细胞结合,并促进神经生长。鼠LNα1链上的RGD序列,可与αvβ3整合素结合。现已发现7种LN分子,8种亚单位(α1,α2,α3,β1,β2,β3,γ1,γ2),与FN不同的是,这8种亚单位分别由8个结构基因编码。徐州开封细胞外基质胶

构成细胞外基质的大分子:Ⅰ型胶原的原纤维平行排列成较粗大的束,成为光镜下可见的胶原纤维,抗张强度超过钢筋。其三股螺旋由二条α1(Ⅰ)链及一条α2(Ⅰ)链构成。每条α链约含1050个氨基酸残基,由重复的Gly-X-Y序列构成。X常为Pro(脯氨酸),Y常为羟脯氨酸或羟赖氨酸残基。重复的Gly-X-Y序列使α链卷曲为左手螺旋,每圈含3个氨基酸残基。三股这样的螺旋再相互盘绕成右手超螺旋,即原胶原。原胶原分子间通过侧向共价交联,相互呈阶梯式有序排列聚合成直径50~200nm、长150nm至数微米的原纤维,在电镜下可见间隔67nm的横纹。胶原原纤维中的交联键是由侧向相邻的赖氨酸或羟赖氨酸残基氧化后所产生...
- 北京正规细胞外基质胶销售厂家 2025-07-03
- 珠海正规细胞外基质胶服务电话 2025-07-03
- 南京成都细胞外基质胶 2025-07-03
- 北京细胞外基质胶厂家直销 2025-07-03
- 济南正规细胞外基质胶厂家供应 2025-07-03
- 杭州正规细胞外基质胶厂家 2025-07-03
- 石家庄细胞外基质胶厂家现货 2025-07-03
- 上海正规细胞外基质胶报价 2025-07-03
- 珠海细胞外基质胶报价 2025-07-03
- 济南正规细胞外基质胶平均价格 2025-07-03

- 无锡正规细胞外基质胶直销价 2025-07-02
- 宁波细胞外基质胶厂家供应 2025-07-02
- 厦门正规细胞外基质胶厂家推荐 2025-07-02
- 厦门正规细胞外基质胶厂家供应 2025-07-02
- 北京正规细胞外基质胶平均价格 2025-07-02
- 太原细胞外基质胶厂家现货 2025-07-02


- 北京正规细胞外基质胶销售厂家 07-03
- 珠海正规细胞外基质胶服务电话 07-03
- 南京成都细胞外基质胶 07-03
- 深圳外泌体提取试剂推荐厂家 07-03
- 昆明正规外泌体提取试剂厂家供应 07-03
- 北京细胞外基质胶厂家直销 07-03
- 济南正规细胞外基质胶厂家供应 07-03
- 杭州正规细胞外基质胶厂家 07-03
- 广州外泌体提取试剂厂家直销 07-03
- 石家庄细胞外基质胶厂家现货 07-03