- 产地
- 苏州
- 品牌
- 细胞外基质胶
- 型号
- 齐全
- 是否定制
- 是
什么是细胞外基质:多细胞有机体中,细胞周围由多种大分子组成bai的复杂网络,称作细胞外基质。研究表明,细胞外基质并非像过去认为的起惰性支持物的作用,或将细胞连接在一起,形成组织、部位。而是含有大量信号分子,积极参与控制细胞的生长,极性,形状、迁移和代谢活动。对人类细胞的研究表明,细胞外基质中的纤粘蛋白主要由成纤维细胞、上皮细胞等分泌并附着在细胞表面,其作用是促进细胞对基质的贴附,细胞之间的粘着,细胞内微丝及应力纤维的构建。现已经观察到转化的体外培养的成纤维细胞,表面纤维蛋白量减少,与此相关地细胞形态变圆,与培养基底贴附松弛,胞内应力纤维较大减少,细胞密集,重叠生长。这种转化细胞接种入正常机体,常能长成块,并侵润正常组织,发生普遍转移。什么是细胞外基质:多细胞有机体中,细胞周围由多种大分子组成的复杂网络,称作细胞外基质。杭州太原细胞外基质胶
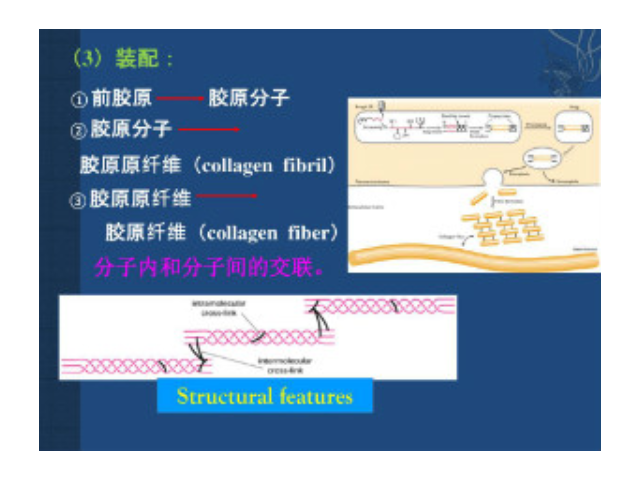
构成细胞外基质的大分子:Ⅰ型胶原的原纤维平行排列成较粗大的束,成为光镜下可见的胶原纤维,抗张强度超过钢筋。其三股螺旋由二条α1(Ⅰ)链及一条α2(Ⅰ)链构成。每条α链约含1050个氨基酸残基,由重复的Gly-X-Y序列构成。X常为Pro(脯氨酸),Y常为羟脯氨酸或羟赖氨酸残基。重复的Gly-X-Y序列使α链卷曲为左手螺旋,每圈含3个氨基酸残基。三股这样的螺旋再相互盘绕成右手超螺旋,即原胶原。原胶原分子间通过侧向共价交联,相互呈阶梯式有序排列聚合成直径50~200nm、长150nm至数微米的原纤维,在电镜下可见间隔67nm的横纹。胶原原纤维中的交联键是由侧向相邻的赖氨酸或羟赖氨酸残基氧化后所产生的两个醛基间进行缩合而形成的。北京正规细胞外基质胶供应商细胞外基质中不同的机械特性对细胞行为和基因表达都有影响。

细胞外基质:在生物学中,细胞外基质是细胞外大分子(如胶原蛋白、酶和糖蛋白)组成的三维网络,为周围细胞提供结构和生化支持。因为在不同的多细胞谱系中,其多细胞化特性是自立进化的,因此,细胞外基质的组成在不同的多细胞结构之间有所不同;然而,细胞粘附、细胞间通讯和分化是细胞外基质的共同功能。动物细胞外基质包括间质基质和基底膜。间质基质存在于各种动物细胞之间(即细胞间隙中)。多糖和纤维蛋白的胶状物填充了细胞间隙,并作为压缩缓冲液,抵御施加在细胞外基质上的压力。基底膜是细胞外基质的片状沉积物,其上有各种上皮细胞。
单个的蛋白聚糖和透明质酸-蛋白聚糖复合物直接与胶原纤维连接形成动物细胞外的纤维-网络(fiber-network)结构,不同类型的胶原和不同类型的蛋白聚糖连接形成不同的纤维-网络,对于提高细胞外基质的连贯性起关键作用。此外,蛋白聚糖还可作为细胞粘着的暂时性或长久性的位点。蛋白聚糖在细胞外基质中的功能是什么:蛋白聚糖的这种性质,使细胞表面具有较大的可塑性,从而具有抗挤压能力,对细胞起保护作用。由于透明质酸以可溶的形式游离存在,所以在细胞外体液和滑液(synovialfluid)中透明质酸的浓度很高,其结果提高了体液和滑液的粘度和润滑性。纤连蛋白与细胞外基质大分子结合,促进它们与跨膜整合素的结合。

细胞外基质成分居然能调节葡萄糖代谢过程:确认ECM组分透明质酸与新陈代谢之间的机制联系,用透明质酸酶处理细胞和异种移植物引发糖酵解的强烈增加。这主要通过快速受体酪氨酸激酶介导的mRNA衰变因子ZFP36的诱导来实现,其靶向TXNIP转录物以降解。因为TXNIP促进葡萄糖转运蛋白GLUT1的内化,其急剧下降使质膜上的GLUT1富集。在功能上,需要通过透明质酸酶诱导糖酵解,同时加速细胞迁移。ECM重塑和代谢之间的这种相互关联在动态组织状态中表现出来,包括部位发生和胚胎发生。研究人员提出ECM重塑作为急性细胞-外在代谢调节的附加节点,可以直接读出周围组织的结构状态并相应地增强细胞行为。细胞外基质调节细胞的动态行为。此外,它能隔离多种细胞生长因子。长沙细胞外基质胶
细胞外基质并非像过去认为的光光起惰性支持物的作用,或将细胞连接在一起,形成组织、部位。杭州太原细胞外基质胶
根据胶原的结构和功能可将其分为:网状胶原(networkformingcollagen)如Ⅳ、Ⅷ和Ⅹ型胶原,主要分布于基底膜中。与纤维性胶原不同,其端肽不被去除。两条Ⅳ型前胶原肽链的羧基端肽(NC1)端-端相连形成二聚体,四条前胶原肽链的氨基端肽(7S)端-端形成四聚体,从而相互交联成三维网状结构。在肝脏中,Ⅳ型胶原主要分布于血管和胆管的基底层,而且还分布于汇管区的成纤维细胞周围及正常肝血窦的Disse腔中。Ⅷ型胶原常与弹性纤维一起分布于肝脏的汇管区和包膜中,其功能尚不清楚。杭州太原细胞外基质胶

细胞外基质层粘连蛋白:(laminin,LN)LN也是一种大型的糖蛋白,与Ⅳ型胶原一起构成基膜,是胚胎发育中出现较早的细胞外基质成分。LN分子由一条重链(α)和二条轻链(β、γ)借二硫键交联而成,外形呈十字形,三条短臂各由三条肽链的N端序列构成。每一短臂包括二个球区及二个短杆区,长臂也由杆区及球区构成。LN分子中至少存在8个与细胞结合的位点。例如,在长臂靠近球区的。链上有IKVAV五肽序列可与神经细胞结合,并促进神经生长。鼠LNα1链上的RGD序列,可与αvβ3整合素结合。现已发现7种LN分子,8种亚单位(α1,α2,α3,β1,β2,β3,γ1,γ2),与FN不同的是,这8种亚单位分别由8个...
- 长沙正规细胞外基质胶单价 2025-07-10
- 深圳正规细胞外基质胶进货价 2025-07-10
- 合肥正规细胞外基质胶厂家供应 2025-07-10
- 石家庄正规细胞外基质胶直销厂家 2025-07-10
- 芜湖细胞外基质胶推荐厂家 2025-07-09
- 上海正规细胞外基质胶哪家好 2025-07-09
- 成都细胞外基质胶单价 2025-07-09
- 正规细胞外基质胶厂家直销 2025-07-09
- 青岛正规细胞外基质胶厂家直销 2025-07-09
- 合肥正规细胞外基质胶销售厂家 2025-07-09



- 珠海正规细胞外基质胶哪家好 2025-07-09
- 石家庄细胞外基质胶直销厂家 2025-07-09
- 重庆细胞外基质胶生产厂家 2025-07-09
- 宁波正规细胞外基质胶价格 2025-07-09
- 北京正规细胞外基质胶厂家直销 2025-07-09
- 青岛正规细胞外基质胶平均价格 2025-07-09




- 杭州外泌体提取试剂销售厂家 07-10
- 合肥正规细胞外基质胶厂家供应 07-10
- 石家庄正规细胞外基质胶直销厂家 07-10
- 芜湖细胞外基质胶推荐厂家 07-09
- 天津正规外泌体提取试剂厂家批发价 07-09
- 上海正规细胞外基质胶哪家好 07-09
- 成都细胞外基质胶单价 07-09
- 济南正规外泌体提取试剂供应商 07-09
- 正规细胞外基质胶厂家直销 07-09
- 温州正规外泌体提取试剂报价 07-09