- 产地
- 苏州
- 品牌
- 细胞外基质胶
- 型号
- 齐全
- 是否定制
- 是
细胞外基质生理学功能:刚度和弹性:从柔软的脑组织到坚硬的骨组织,细胞外基质可以以不同程度的刚度和弹性存在。细胞外基质的弹性可以相差几个数量级。这一特性主要取决于胶原蛋白和弹性蛋白的浓度,较近已证明细胞外基质的刚度和弹性这个特性在调节许多细胞功能中发挥着重要作用。细胞可以通过施力和测量产生的反作用力来感知环境的机械特性。这发挥着重要作用,因为它有助于调节许多重要的细胞过程,包括细胞收缩,细胞迁移,细胞增殖,分化[和细胞死亡(凋亡)。克制非肌肉肌球蛋白ⅱ阻断了大部分这些效应,表明它们确实与细胞外基质的机械特性有关,这在过去十年中已成为研究的新焦点。植物细胞的细胞壁相当于植物体中的细胞外基质。青岛正规细胞外基质胶直销厂家
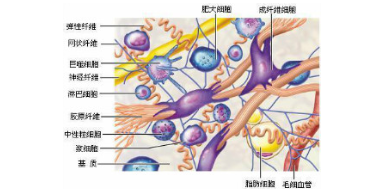
细胞外基质的主要类型及功能:a.骨的胞外基质表现为刚硬的特点,以满足支撑的作用b.软骨是另一种结缔组织,其胞外基质具有一定的韧性c.眼角膜中胞外基质是透明的保护层。如前所述,动物组织的构建既是多细胞相互作用的结果,也是细胞与胞外基质相互作用和接触的结果。胞外基质不光为组织的构建提供的支撑框架,还对与其接触的细胞的存活、分化、迁移、增殖与形态以及其他功能产生重要的调控作用。鉴于细胞外间质的多样性,细胞外间质有多方面的功能。例如,为细胞提供支持和固定、提供组织间的分离方法、调节细胞间的沟通。细胞外间质调节细胞的动态行为。长沙正规细胞外基质胶供应商细胞具有不同的机制以感知和响应外在代谢信号。

细胞外基质的作用:由于细胞外基质对细胞的形状、结构、功能、存活、增殖、分化、迁移等一切生命现象具有很全的影响,因而无论在胚胎发育的形态发生、部位形成过程中,或在维持成体结构与功能完善(包括免疫应答及创伤修复等)的一切生理活动中均具有不可忽视的重要作用。ECM与肾脏纤维化:各种原发性和/或继发性致病原因所导致ECM合成与降解的动态失衡,促使大量ECM积聚而沉积于肾小球、肾间质内,导致肾脏各级血管堵塞,混乱分隔形成肾脏组织形态学改变,较终导致肾单位丧失,肾功能衰竭,进一步发展成为不可逆转的肾小球硬化
细胞外基质是动物组织的一部分,不属于任何细胞。它决定结缔组织的特性,为细胞的生存及活动提供适宜的场所,并通过信号转导系统影响细胞的形状、代谢、功能、迁移、增殖和分化。一旦它们召集到正确的细胞,基质便会展现它的另一大作用:它能依据细胞在基质内承受的张力,引导它们转化成骨骼细胞、肌肉细胞或脂肪细胞。这个张力是每日肌肉运动力量的附加产物。在实验室中,张力则是由人工操作基质硬度来完成。例如,高张力能够使所有进入的干细胞变成肌肉或骨骼,而在相对松弛的基质中,干细胞则会变成脂肪细胞。细胞外基质重建你的身体:细胞外基质(extracellularmatrix,ECM)是由细胞合成并分泌到胞外、分布在细胞表面或细胞之间的大分子,主要是一些多糖和蛋白,或蛋白聚糖。这些物质构成复杂的网架结构,支持并连接组织结构、调节组织的发生和细胞的生理活动。纤连蛋白与细胞外基质大分子结合,促进它们与跨膜整合素的结合。

细胞外基质生理学功能:对基因表达的影响:细胞外基质中不同的机械特性对细胞行为和基因表达都有影响。尽管实现这一点的机制尚未完全解释清楚,但粘附复合物和肌动蛋白-肌球蛋白细胞骨架(其收缩力通过跨细胞结构传递)被认为在尚未发现的分子途径中起着关键作用。对分化的影响:细胞外基质弹性可以指导细胞分化,即细胞从一种细胞类型转变为另一种细胞类型的过程。特别是,原始间充质干细胞(MSCs)已被证明能明确谱系并表现出对组织水平弹性极其敏感的表型。将原始间充质干细胞(MSCs)置于模拟大脑分化成神经元样细胞的柔软基质上,表现出相似的形状、RNAi图谱、细胞骨架标记和转录因子水平。类似地,模拟肌肉的刚度基质是肌源性的,而模拟胶原骨的刚性基质是成骨性的。成肌细胞在纤粘连蛋白上增殖并保持未分化的表型。芜湖成都细胞外基质胶
细胞外基质的生物学作用:调节细胞的增殖。青岛正规细胞外基质胶直销厂家
细胞外基质粘弹性影响细胞行为:组织和细胞外基质的复杂力学行为,讨论了细胞外基质粘弹性对细胞的影响,并描述了粘弹性生物材料在再生医学中的潜在应用。生物材料设计历来没有考虑到粘弹性的重要性,但是展望未来,粘弹性很可能成为许多应用中的关键技术规范(如图5)。粘弹性在调节可能包括多能干细胞、组织驻留干细胞和分化细胞以及免疫细胞在内的各种细胞类型生物学调节中的作用,以便合理设计能够促进组织再生的材料。生物材料的设计也可能需要将细胞感知的局部粘弹性特性与实现再生或工程组织机械稳定性所需的更大的组织尺度特性相分离。因此,具有可控粘弹性的生物材料的出现可能会改变生物材料在再生医学中的应用。青岛正规细胞外基质胶直销厂家

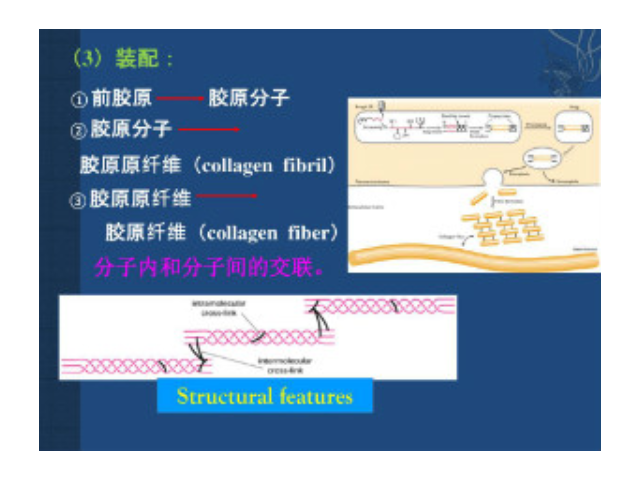
构成细胞外基质的大分子:Ⅰ型胶原的原纤维平行排列成较粗大的束,成为光镜下可见的胶原纤维,抗张强度超过钢筋。其三股螺旋由二条α1(Ⅰ)链及一条α2(Ⅰ)链构成。每条α链约含1050个氨基酸残基,由重复的Gly-X-Y序列构成。X常为Pro(脯氨酸),Y常为羟脯氨酸或羟赖氨酸残基。重复的Gly-X-Y序列使α链卷曲为左手螺旋,每圈含3个氨基酸残基。三股这样的螺旋再相互盘绕成右手超螺旋,即原胶原。原胶原分子间通过侧向共价交联,相互呈阶梯式有序排列聚合成直径50~200nm、长150nm至数微米的原纤维,在电镜下可见间隔67nm的横纹。胶原原纤维中的交联键是由侧向相邻的赖氨酸或羟赖氨酸残基氧化后所产生...
- 北京正规细胞外基质胶销售厂家 2025-07-03
- 上海正规细胞外基质胶报价 2025-07-03
- 济南正规细胞外基质胶平均价格 2025-07-03
- 细胞外基质胶销售厂家 2025-07-03
- 苏州正规细胞外基质胶厂家现货 2025-07-02
- 无锡正规细胞外基质胶直销价 2025-07-02
- 宁波细胞外基质胶厂家供应 2025-07-02
- 厦门正规细胞外基质胶厂家推荐 2025-07-02
- 厦门正规细胞外基质胶厂家供应 2025-07-02
- 北京正规细胞外基质胶平均价格 2025-07-02



- 深圳细胞外基质胶产品介绍 2025-07-02
- 芜湖细胞外基质胶厂家现货 2025-07-02
- 珠海细胞外基质胶直销价 2025-07-02
- 徐州细胞外基质胶厂家现货 2025-07-02
- 厦门正规细胞外基质胶厂家现货 2025-07-01
- 南京正规细胞外基质胶报价 2025-07-01

- 济南正规细胞外基质胶平均价格 07-03
- 细胞外基质胶销售厂家 07-03
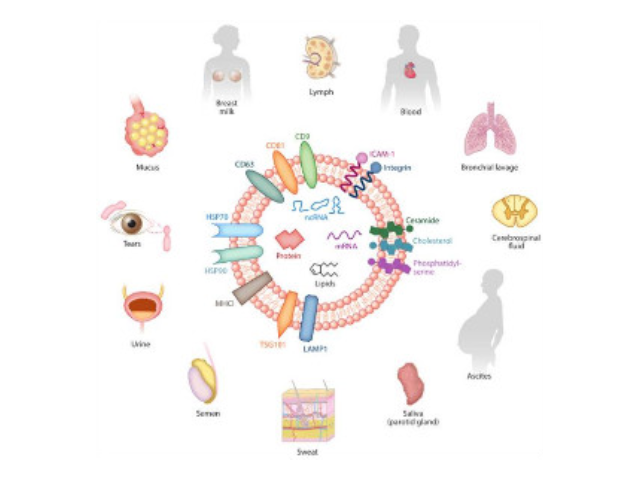
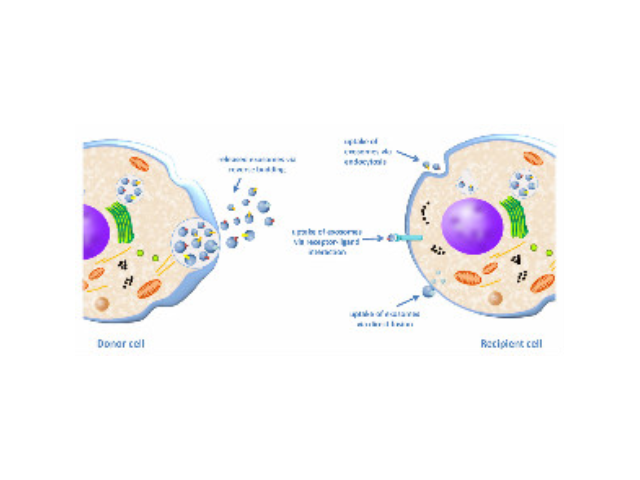
- 唐山外泌体提取试剂直销价 07-02
- 珠海正规外泌体提取试剂供应商 07-02
- 苏州正规细胞外基质胶厂家现货 07-02
- 芜湖正规外泌体提取试剂价格 07-02
- 长沙正规外泌体提取试剂 07-02
- 重庆外泌体提取试剂推荐厂家 07-02
- 无锡正规细胞外基质胶直销价 07-02
- 芜湖正规外泌体提取试剂厂家推荐 07-02